邮箱地址

前沿资讯
发布时间:2023-10-23 09:23:28 作者:青岛海大海洋寡糖科技有限公司 来源:本站
文章题目:Synthetic phosphoethanolamine-modified oligosaccharides reveal the importance of glycan length and substitution in biofilm-inspired assemblies
发表期刊:Nature Communications
影响因子:17.694(2022)
通讯单位:Department of Biomolecular Systems

细菌生物膜基质是蛋白质和多糖的纳米复合材料,具有显着的机械性能。大肠杆菌(E.coli)生物膜的细胞外基质(ECM)的主要成分是卷曲纤维——细菌功能性淀粉样蛋白和纤维素(图1-a)。理解和调整蛋白质成分的研究已经比较广泛,而多糖部分仍被忽视。近期的研究发现,某些细菌(如大肠杆菌和肠沙门氏菌)能够产生带有磷酸乙醇胺(pEtN)取代基的化学改性纤维素,这种卷曲的且带有pEtN 取代基的纤维素复合物产生的生物膜具有较强的弹性和对膀胱上皮细胞的粘附性。这表明,碳水化合物可以调节生物膜的特性,并且可能是定制纤维素材料用于组织工程、生物技术和食品工业的基础。
因此,本篇文章通过自动聚糖组装(AGA)合成的九种pEtN纤维素寡糖的合成,这些寡糖具有不同的链长、程度和 pEtN 取代的模式。研究了这些聚糖与curli (R5)的代表性淀粉肽的相互作用。
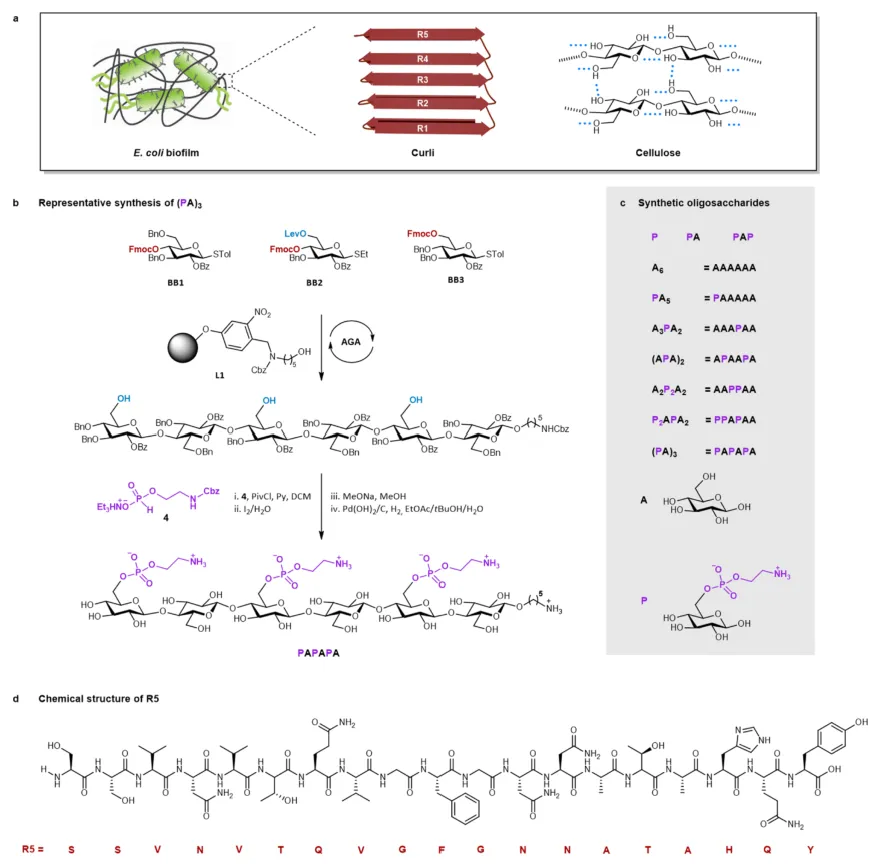
图1 大肠杆菌生物膜及其代表性基质成分的合成
a.大肠杆菌生物膜的卡通表示;b.pEtN六糖的代表性合成,AGA包括糖基化、加帽和Fmoc去保护的循环,最终Lev去保护释放了在AGA 后步骤中官能化的羟基,AGA的反应条件在SI中报告,PivCl=新戊酰氯,py =吡啶,MeONa =甲醇钠;c.本工作中合成的寡糖集合;d.肽R5的化学结构。
pEtN 取代的寡糖通过AGA和AGA后步骤的组合制备(图1-b)。纤维素骨架由AGA构建,在固体支持物L1上进行糖基化和 Fmoc去保护循环。BB1和BB2被战略性地组装以产生具有所需羟基模式的低聚物,BB3用于组装的最后一个循环。Lev去除后,寡糖骨架从固体支持物上切割下来并进行AGA后转化。可用的羟基与H-膦酸酯4偶联,在用碘水溶液 30 氧化后得到受保护的磷酸化化合物。最终制备了九种两性离子化合物:带有一个 pEtN 基团的单糖和二糖,带有两个pEtN基团的三糖,以及被一个、两个或三个pEtN单元取代的六个六糖(图1-c)。
文章选择了curli28的CsgA单元中淀粉样蛋白生成最多的重复的合成分子(R5,如图1-d)作为生物膜组装的人红模型。随后,在各自的寡糖存在下重复相同的组装过程。
首先,筛选了寡糖长度对R5聚集的影响,结果发现,与较短的类似物P、PA和PAP的共组装不会显着影响R5的结构转变速率,而在存在较长的六糖的情况下,R5向β折叠构象的二级结构转变较慢(图2-a),能够检测到明确的纤维状结构,并且发现,纤维随着寡糖链的长度而变长。
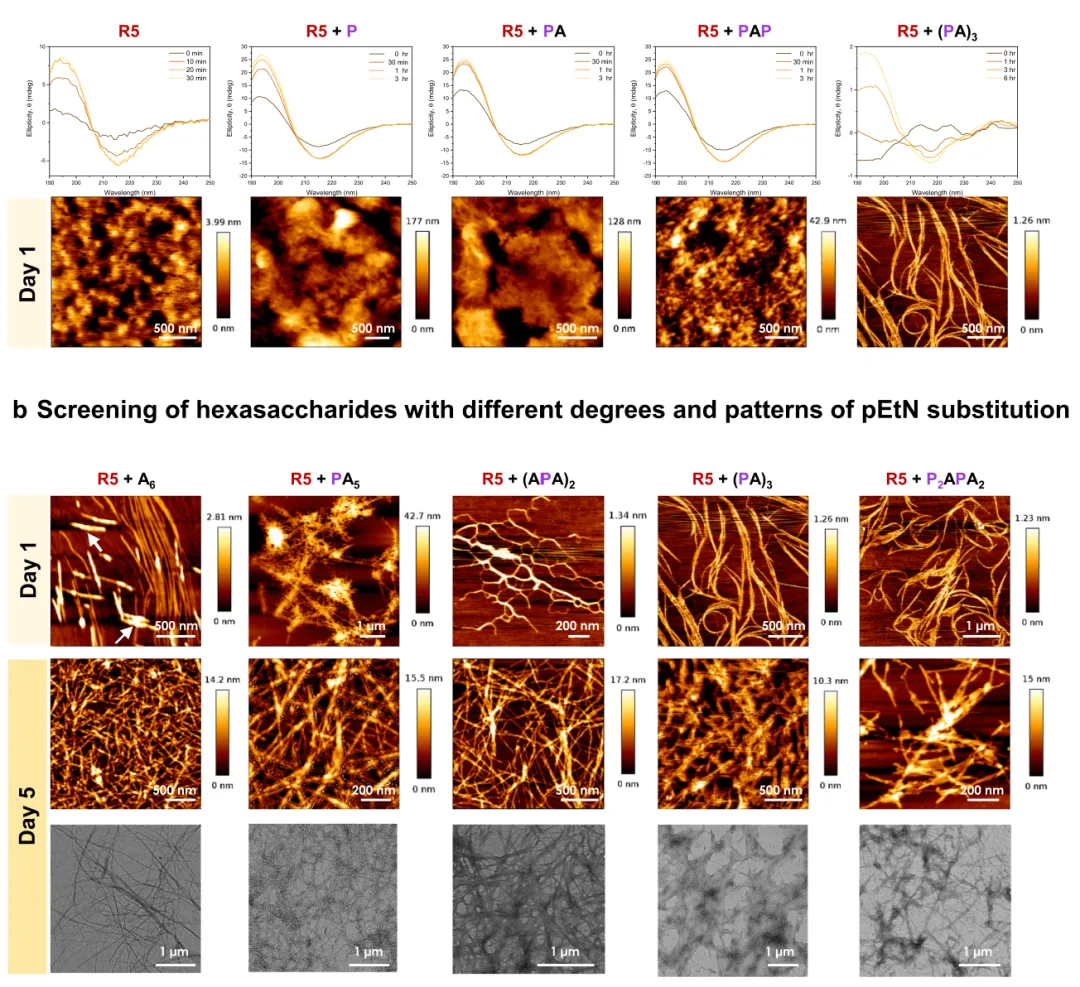
图2 在选定的寡糖存在下组装R5
a. 筛选不同长度的寡糖。单独含有R5或存在不同寡糖的样品的CD光谱和AFM(第1天)。单独的R5样品会导致不明确的聚集,而当R5在寡糖存在下组装时,会产生不同尺寸的纤维状结构。b.筛选具有不同程度和模式的 pEtN 取代的六糖。AFM(第1天和第5天)和TEM(第5天)样品在存在具有不同程度和pEtN取代模式的六糖的情况下的R5。A6的聚合用白色箭头表示。
接下来,文章对七种纤维素六糖在R5的组装上具有不同程度和模式的pEtN取代进行了检测。pEtN取代的差异影响了R5的二级结构转变速率,在(PA)3存在下制备的样品转变最慢(图2-a)。另外,含有未取代的纤维素低聚物A6的R5样品组装成为细纤维,在5天内变成纤维网络,虽然含有三取代低聚物((PA)3和P2APA2)的样品在第1天已经显示出长而明确的原纤维,但取代较少的类似物形成较短的聚集体,有趣的是,对R5和 (PA)3观察到的纤维均为经典的卷曲形状,在第5天,所有样品都形成了纤维网络(图2-b)。
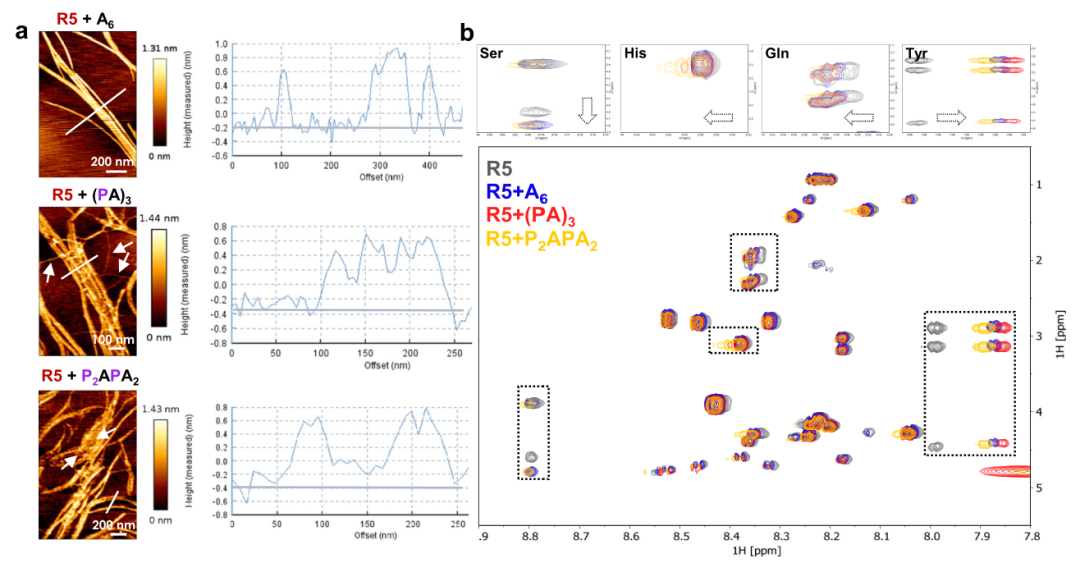
图3 在存在三种不同寡糖的情况下,R5 组装产生的纤维的结构分析。
a.第1天纤维的 AFM 图像。所有三个样品的平均纤维高度约为 0.8 nm。啮合矩阵用白色箭头突出显示。b.仅包含 R5(灰色)和存在 A6(蓝色)、(PA)3(红色)和 P2APA2(黄色)的样品的1H-1H TOCSY 光谱的选定区域的叠加。在25 °C下,在H2O/D2O (9:1)中以200 µM 的R5浓度记录每个光谱。突出显示主要受寡糖存在影响的四个酰胺质子(上图),显示化学位移(His、Gln、Tyr)或信号展宽(Ser)的变化。
在存在A6、(PA)3或P2APA2的情况下,从R5样品中获得的纤维显示出相似的z-高度,约为0.8 nm(图3-a),这表明原纤维建立在相同的肽核心上。相比之下,由含有R5和(PA)3或P2APA2的样品产生的纤维嵌入薄基质中。未染色的TEM图像证实了原纤维周围存在基质,显示纤维比周围更亮(图2-b)。
含有R5和A6、(PA)3和P2APA2的三个样品分别显示出所选氨基酸的更高溶解度和化学位移扰动(图3-b)。酪氨酸、谷氨酰胺、组氨酸和丝氨酸是所有三个样品中受影响最大的氨基酸。总之,寡糖的存在减缓了R5向β折叠构象的转变,有利于形成长淀粉样蛋白纤维而不是不明确的聚集体。这可能是直接的肽-寡糖相互作用导致离子型 pEtN 基团暴露于水或由于寡糖的存在导致肽环境发生变化的结果。
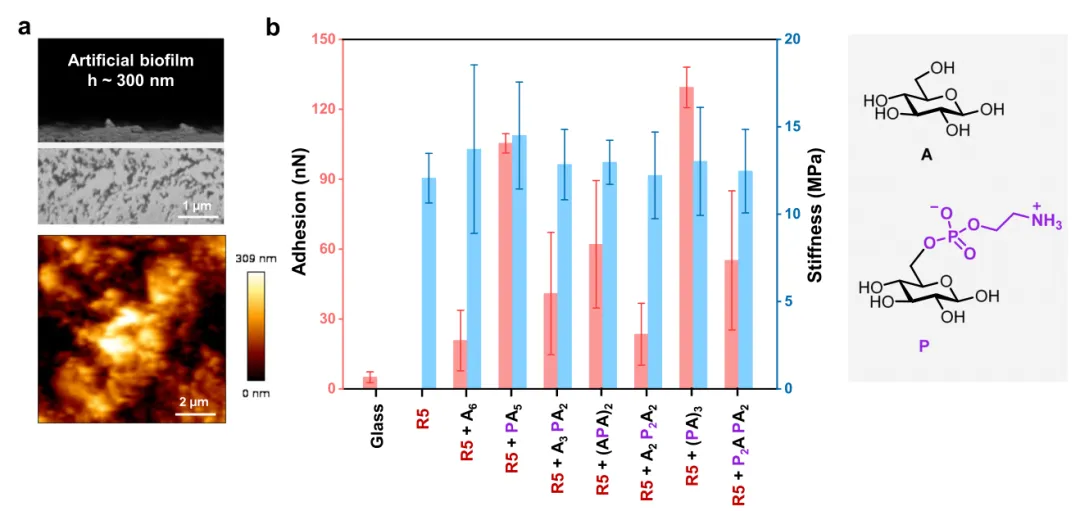
图4 在不同六糖存在下由R5组装产生的人工基质的机械性能。
a .R5与(PA)3的薄膜的横截面SEM和AFM图像。b.由AFM力-距离曲线产生的附着力和刚度。每个数据点对应于50-100力测量的平均值,误差条代表平均值的标准偏差。由于薄膜的不均匀性,仅R5的附着力无法测量。
接下来对该人造生物膜的机械性能做了探索。将产生纤维的共同组装样品滴铸在玻璃载玻片上,以制备厚度约为300 nm的人造生物膜基质。使用AFM力-距离曲线分析探索其化学特性(图4)。结果发现,肽纤维是人造基质的主要结构成分,pEtN修饰的寡糖的存在显着增强了粘附性。紧邻的多个pEtN取代基(例如A2P2A2)导致低得多的粘附力,强调了取代模式在确定薄膜机械性能中的重要性。
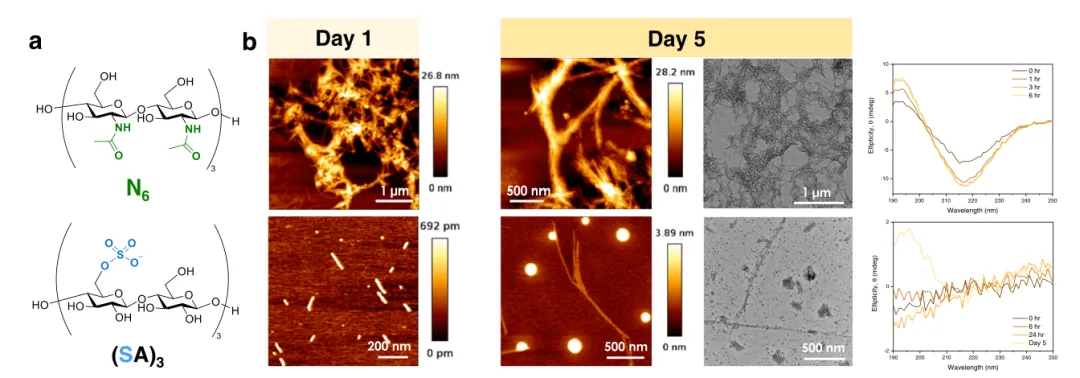
图5 探索非天然寡糖对 R5 组装的影响。
a .N-乙酰氨基葡萄糖六糖 N6和硫酸化六糖 (SA)3 的化学结构。b. AFM(第1天和第5天)、TEM(第5天)图像和CD光谱在N6或(SA)3存在下用 R5 制备的原纤维。
天然修饰的pEtN纤维素的发现为在碳水化合物成分的工程化过程中生成可调节材料开辟了机会。为了探索不同聚糖修饰对R5聚集的影响,按照既定方案制备了天然细菌生物膜中不存在的两种六糖(图5-a)。在N-乙酰氨基葡萄糖六糖N6的存在下,R5向β折叠的二级结构转变在不到3小时内完成,形成了进一步聚集成超分子束的小于1 µm的原纤维(图5-b)。相反,带负电荷的硫酸化六糖(SA)3 中断了R5向β折叠构象的转变和原纤维的形成(图5-b)。这种抑制可能是带负电荷的寡糖(SA)3和R5上的阳离子基团之间强烈的共轭相互作用的结果,强调了两性离子pEtN基团在指导R5聚集中的重要性。硫酸化六糖(SA)3抑制淀粉样蛋白形成的能力使该化合物成为治疗神经疾病或作为抗菌剂的新方法的有趣起点。
本文合成了一系列pEtN修饰的寡糖,完全控制了链长、取代程度和模式,为研究复杂的生物系统提供了基本标准。将寡糖与代表 curli的合成肽R5一起孵育,以生成大肠杆菌生物膜ECM的模块化模型并分解其复杂性。虽然较短的寡聚体几乎没有影响,但较长的六糖减慢了二级结构向肽R5的β折叠的转变,从而诱导了延伸纤维结构的生长。低聚糖精细结构显着影响复合材料的纤维生长速率和力学性能。pEtN的取代程度和模式都会影响粘附。相反,所有样品的刚度保持不变,表明其与肽成分的紧密联系。代谢工程和/或定向进化方法可能会在体内引入此类修饰并产生具有非天然修饰的新型纤维素材料。
原文链接:
https://doi.org/10.1038/s41467-022-31633-5
作者:王维雯
审核:李全才,吕友晶
编辑:邵萌
如有侵权,请联系删除
